2021年8月,上海师范大学杨仲南教授团队与黄学辉教授课题组合作,于Molecular Plant 在线发表了题为 Non-cell Autonomous Cell Wall Transition Defects Mediate Thermo/photoperiod-sensitive Genic Male Sterility 的研究论文,揭示植物光温敏育性转换的细胞学机制。

自上世纪80年代发现水稻光温敏不育系以来,光温敏不育系已经广泛应用于水稻杂交制种。然而,温度和光照作为共同的环境信号,是如何恢复不同植物不同遗传位点决定的不育系,尚不清楚。杨仲南团队在前期工作中通过拟南芥光温敏不育突变体reversible male sterile (rvms)的研究,揭示了低温或短光照条件下的缓慢发育是植物光温敏育性转换的一个共性机制(Zhu et al., Nat. Plants 6:360-367; Zhang et al., Plant Physiol 184:923-932)。
植物细胞受到细胞壁的保护。减数分裂完成后形成的小孢子包裹在四分体壁(由果胶壁和胼胝质壁组成)中。随着小孢子发育成花粉粒,四分体壁降解,被花粉外壁取代,这个过程被称为小孢子壁的转换期。在这个工作中,研究人员筛选到了一个rvms突变体的恢复系(res2),该突变体能够在常温下恢复rvms的育性。该基因编码一个已报道的负责四分体果胶壁降解的多聚半乳糖醛酸酶QUARTET 3(QRT3)。此前该实验室发现,绒毡层MYB转录因子MS188直接调控孢粉素合成途径基因的表达,负责花粉外壁的合成(Wang et al., Plant Physiol 178: 283-294)。他们发现MS188也直接调控RES2/QRT3的表达。因此,在小孢子发育成成熟花粉过程中,其细胞壁的转换受到绒毡层细胞MS188主导的非自主途径控制(图1)。
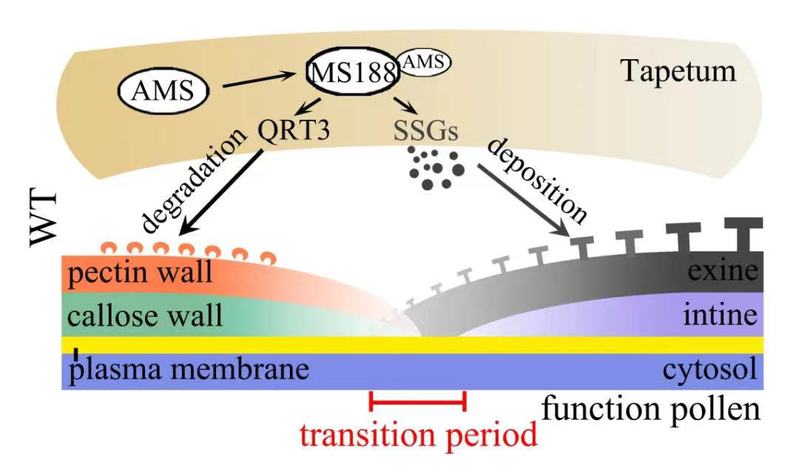
图1. 绒毡层转录因子MS188调控果胶壁的降解和花粉外壁的合成
目前报道的拟南芥光温敏基因都直接或间接与花粉壁形成有关,相关的突变体花粉壁的缺陷无法保护小孢子渡过转换期而破裂,呈现雄性不育表型(图2)。
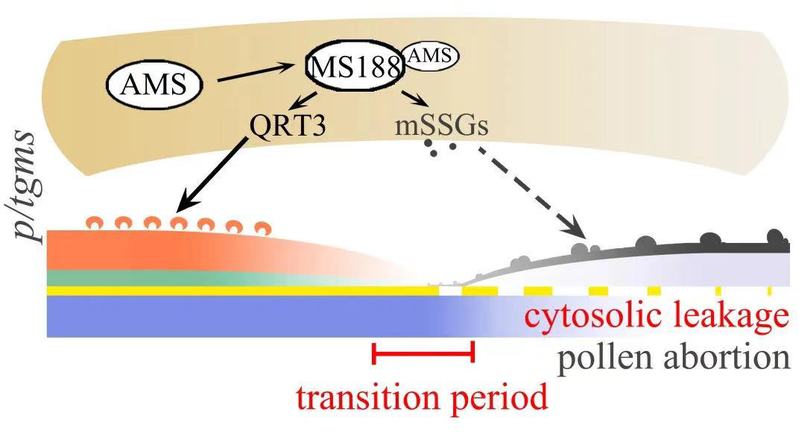
图2. 光温敏突变体中外壁缺陷的小孢子无法渡过转换期而破裂降解
除了RES2/QRT3外,果胶甲基酯酶QRT1以及多聚半乳糖醛酸酶QRT2也参与四分体果胶壁的降解。这些qrt系列突变体的果胶壁降解延迟,都能不同程度恢复rvms及其他P/TGMS株系的育性。与之相反的是,如果提前表达RES2基因会导致果胶壁过早降解,抵消了低温对rvms育性恢复的效果。延迟降解的果胶壁可能提供额外的保护作用,帮助P/TGMS小孢子发育成有功能的花粉,从而恢复育性(图3)。
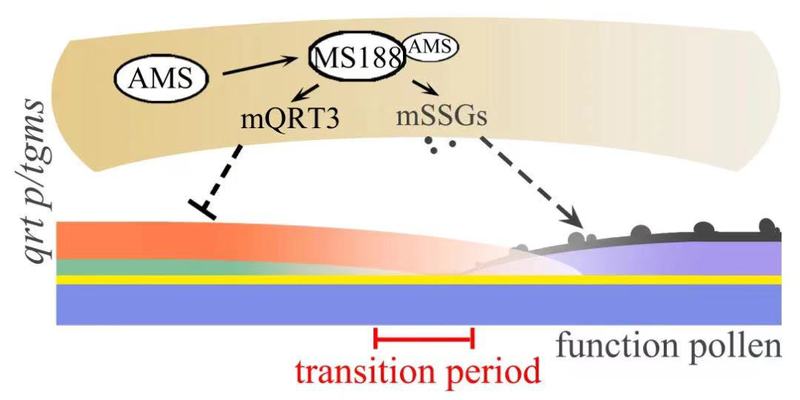
图3. 延迟降解的果胶壁为外壁缺陷的小孢子提供了额外保护从而恢复光温敏植物育性
基于这些研究结果,研究人员提出了光温敏育性恢复的细胞学机制。光温敏植物中缺陷的花粉壁无法满足小孢子发育的需求导致花粉破裂植株雄性不育。各种环境因素包括低温、短光照、弱光等条件下,小孢子发育缓慢,对细胞壁保护作用要求降低,光温敏植物中缺陷的花粉壁能够满足小孢子发育的需求,从而恢复育性(图4)。目前,拟南芥中的光温敏遗传位点以及光温敏恢复机制能否适用于水稻等作物,该课题组仍在研究之中。
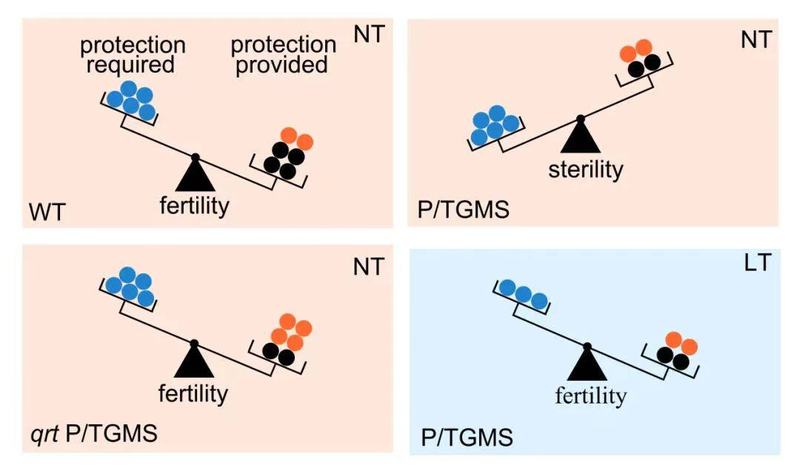
图4. 植物光温敏育性恢复的细胞水平机制
在该研究中,石强胜博士和楼悦老师为论文共同第一作者,杨仲南教授与朱骏教授为共同通讯作者。该研究得到了国家自然科学基金、上海市教委和科委等课题的资助。