光是调控植物生长发育的重要环境信号。植物种子在土壤中发芽后,通过暗形态建成促进幼苗快速伸长,出土见光;然后通过光受体介导的光信号转导促进光形态建成,来抑制下胚轴伸长、促进子叶和叶片的扩展以及叶绿素的合成,使幼苗得以进行光合作用,从而实现自养生长。bHLH类转录因子PIFs(包括PIF1、PIF3、PIF4和PIF5)是红光/远红光受体光敏色素(phytochromes)的关键下游因子,通过促进下胚轴的伸长来抑制光形态建成过程。红光下激活的光敏色素B (phyB)通过与PIFs互作,诱导其磷酸化和蛋白降解,从而促进光形态建成。H2A.Z作为一种组蛋白变体,在植物温度感知、发育和开花时间调控等过程中发挥重要作用;SWR1复合体则负责催化H2A.Z在染色质中的占位。目前为止,已知PIF4和SWR1复合体在调控温度依赖的植物生长过程中具有十分紧密的关系,但是二者发挥作用的生化分子机制尚不明确。
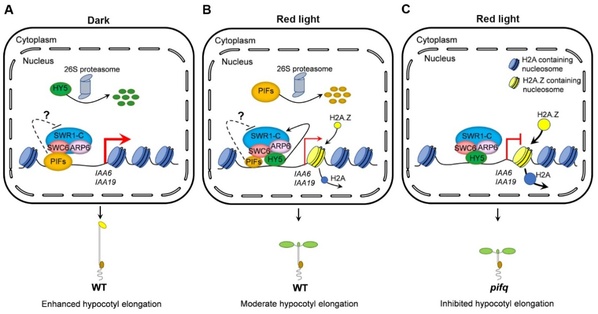
2023年4月28日,Journal of Genetics and Genomics在线发表生命科学学院杨洪全教授和茅志磊副研究员团队题为“PIFs interact with SWC6 to regulate H2A.Z deposition andphotomorphogenesis in Arabidopsis”的研究论文。该研究发现拟南芥光敏色素互作因子PIFs通过与SWR1复合体亚基SWC6直接互作,抑制SWC6与光形态建成关键正调控因子HY5的结合,减少H2A.Z在生长素响应基因上的占位,从而抑制光形态建成。

该研究首先发现在酵母细胞和体外系统中PIF1、PIF3、PIF4和PIF5分别与SWC6存在互作;在烟草细胞共表达PIF1、PIF3、PIF4和PIF5分别与SWC6的两两组合,在细胞核中可定位到相同的核斑点;免疫共沉淀实验显示PIF3和PIF4与SWC6在拟南芥体内存在互作。通过把swc6和arp6分别与pifq四突变体杂交,得到swc6 pifq和arp6 pifq五突变体,比较红光下下胚轴长度,发现SWC6和ARP6在红光下主要通过依赖于PIFs来控制下胚轴的伸长。基因表达结果显示PIFs与SWC6和ARP6拮抗地调控生长素响应基因IAA6、IAA19、 IAA20以及IAA29的表达;ChIP-qPCR结果显示pifq突变体中在IAA6和IAA19基因上的H2A.Z占位水平比WT明显上升。通过蛋白体外和体内实验,发现PIFs可能通过与HY5竞争结合SWC6来抑制SWR1复合体在这些基因上的催化活性,从而调控H2A.Z的占位和光形态建成。
|