2022年7月3日,上海师范大学生命科学学院杨仲南/朱骏教授团队于《Plant Biotechnology Journal》发表了题为“Mutation of GMC oxidoreductase leads to thermosensitive genic male sterility in rice and Arabidopsis”的研究论文,该研究发现了一个新的水稻温敏不育的遗传位点,从细胞学层次解析了温敏不育的机制,反映了水稻和拟南芥温敏不育的共同性和差异性。

自上世纪80年代发现水稻光温敏不育系以来,光温敏不育系已经广泛应用于水稻两系杂交制种。经过四十多年的努力,育种学家发现了十多个水稻光温敏不育系。近年研究发现,生产上应用的水稻两系光温敏不育系中90%以上的品种都是利用现有的tms5位点。一些年份偶发的夏季低温潮导致带有该位点不育系育性部分恢复,严重影响杂交制种的纯度。因此农业生产上迫切需要不育性更为稳定的光温敏不育系。课题组先前对模式植物拟南芥光温敏不育机制开展了系统深入的研究,发现低温下小孢子发育缓慢是育性恢复的共性机制(Zhu et al., 2020, Nature Plants 6:360-367)。光敏不育系育性恢复也是由短光照条件下小孢子发育缓慢所致,因此光照和温度共同通过调控小孢子发育进程从而恢复育性(Zhang et al., 2020, Plant Physiology 184: 923-932)。课题组筛选到一系列光温敏不育系的恢复子,揭示了低温下缓慢发育降低小孢子对细胞壁保护作用的要求从而恢复育性的细胞学机制(Shi et al., 2021, Molecular Plant 14:2104-2114; Wang et al.,2021, Journal of Integrative Plant Biology 64: 717-730)。
课题组对拟南芥光温敏不育机制深入解析的基础上,发明了水稻光温敏不育系的创制技术,获得了多个中花11(ZH11)为背景的光温敏不育系。其中一个命名为ostms18的株系在高温(HT)下(29 ℃)表现为完全不育,但在低温(LT)下(23 ℃)育性正常,且其他农艺性状与野生型相比没有明显区别。在 2021 年上海地区偶发的夏季低温潮的影响下,tms18与同样为中花11背景的tms5突变体相比,其高温不育性状显著好于tms5。通过将该位点引入不同背景的水稻品种中发现,含有其纯合突变位点的植株仍表现为雄性不育,因此该遗传位点在两系杂交水稻育种中具有较好的应用潜力(图1)。
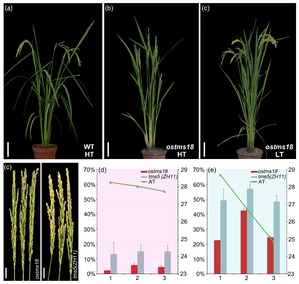
图1. ostms18的温敏不育性状优于同样背景的tms5位点
OsTMS18编码一个在花药中高效表达的葡萄糖-甲醇-胆碱(GMC)氧化还原酶。先前其他课题组报道该基因强等位突变体np1为完全雄性不育。ostms18中该基因第二个外显子一个氨基酸发生突变(Gly-Ser)导致光温敏雄性不育。细胞学分析表明,ostms18 突变体在高温下花粉外壁第二层的结构发生异常,导致花粉破裂,但低温下外壁第二层的结构虽然变薄但较为完整。拟南芥中同源基因AtTMS18 的突变体在常温下(24 ℃)育性正常,高温条件下(28 ℃)育性显著降低,同样表现为温敏不育性状。此外,OsTMS18以及 AtTMS18基因分别受到了绒毡层花粉外壁形成关键转录因子 OsMS188 和 MS188 的直接调控,表明它们都参与到了花粉壁的形成过程中(图2)。
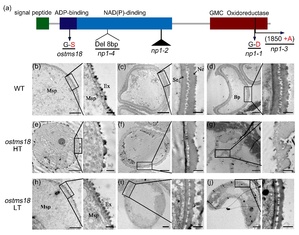
图2. OsTMS18基因结构及野生型和突变体在高低温下的细胞学分析
基于以上研究结果,研究者提出了水稻光温敏不育系ostms18育性恢复的细胞学机制。在小孢子发育成花粉过程中,需要进行细胞壁的转换。四分体壁逐渐降解同时紧随花粉外壁形成,随后在花粉外壁和质膜之间形成的花粉内壁对成熟花粉粒起保护作用。ostms18中花粉外壁缺陷导致花粉破裂;低温条件下的缓慢发育,有缺陷的花粉壁足以保护小孢子继续形成花粉内壁从而恢复育性。另外,该文章对于不同植物的光温敏育性恢复的差异性进行了分析。水稻在夏季强日照和高温条件下生长,花粉形成过程中需要更强的花粉壁保护。而拟南芥作为阴生模式植物,小孢子发育过程中对花粉壁保护的要求不高,相关基因的突变体育性更容易恢复。这些结果有助于对于植物光温敏育性恢复机制的理解(图3)。
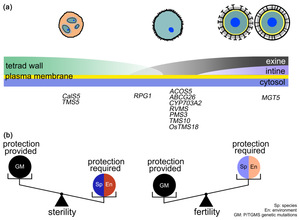
图3. 植物光温敏育性恢复的细胞水平机制
在该研究中,上海师范大学博士生张艳飞和李月灵博士为论文共同第一作者,杨仲南教授与朱骏教授为共同通讯作者,黄学辉教授也参与了部分研究工作。该研究得到了国家自然科学基金、上海市教委和科委等课题的资助。
文章链接:https://onlinelibrary.wiley.com/doi/10.1111/pbi.13886